Taxonomic group: bacteria / Firmicutes
(Phylum: Firmicutes)
The structure was elucidated in this paperNCBI PubMed ID: 28821115Publication DOI: 10.1016/j.carbpol.2017.06.113Journal NLM ID: 8307156Publisher: Elsevier
Correspondence: P. López <plg
cib.csic.es>
Institutions: Centro de Investigaciones Biológicas (CIB), CSIC, Ramiro de Maeztu 9, 28040 Madrid, Spain, Laboratoire de Microbiologie Fondamentale etAppliquée, Faculté des Sciences de la Nature et de la Vie, Université d'Oran 1 Ahmed Ben Bella, Es Senia, 31100 Oran, Algeria, Departamento de Química Aplicada, Facultad de Química, Universidad del País Vasco, 20018 San Sebastián, Spain, Departamento de Microbiología y Bioquímica de Productos Lácteos, Instituto de Productos Lácteos de Asturias (IPLA), CSIC, Paseo Río Linares s/n, 33300 Villaviciosa, Asturias, Spain, Departamento de Biotecnología, Instituto de Agroquímica y Tecnología de Alimentos (IATA), CSIC, Av. Agustín Escardino 7, 46980 Paterna, Spain
Dextrans synthesised by three Leuconostoc mesenteroides strains, isolated from mammalian milks, were studied and compared with dextrans produced by Lc. mesenteroides and Lactobacillus sakei strains isolated from meat products. Size exclusion chromatography coupled with multiangle laser light scattering detection analysis demonstrated that the dextrans have molecular masses between 1.74x10(8)Da and 4.41x10(8)Da. Rheological analysis of aqueous solutions of the polymer revealed that all had a pseudoplastic behaviour under shear conditions and a random, and flexible, coil structure. The dextrans showed at shear zero a difference in viscosity, which increased as the concentration increased. Also, the purified dextrans were able to immunomodulate in vitro human macrophages, partially counteracting the inflammatory effect of Escherichia coli O111:B4 lipopolysaccharide. During prolonged incubation on a solid medium containing sucrose, dextran-producing bacteria showed two distinct phenotypes not related to the genus or species to which they belonged. Colonies of Lc. mesenteroides CM9 from milk and Lb. sakei MN1 from meat formed stable and compact mucoid colonies, whereas the colonies of the other three Leuconostoc strains became diffuse after 72h. This differential behaviour was also observed in the ability of the corresponding strains to bind to Caco-2 cells. Strains forming compact mucoid colonies showed a high level of adhesion when grown in the presence of glucose, which decreased in the presence of sucrose (the condition required for dextran synthesis). However no influence of the carbon source was detected for the adhesion ability of the other Lc. mesenteroides strains, which showed variable levels of binding to the enterocytes.
Lactic acid bacteria, exopolysaccharides, adhesion, Dextran, Leuconostoc mesenteroides, immunomodulation, Rheological properties
Structure type: polymer chemical repeating unit
Location inside paper: p.650, fig.1C
Trivial name: dextran
Compound class: EPS
Contained glycoepitopes: IEDB_142488,IEDB_144998,IEDB_146664,IEDB_158538,IEDB_2346541,IEDB_983931,SB_192
Methods: methylation, IR, sugar analysis, statistical analysis, immunological assays, SEC-MALLS, bacterial adhesion to Caco-2 cells, rheological analysis
NCBI Taxonomy refs (TaxIDs): 1245Reference(s) to other database(s): GTC:G33005VJ
Show glycosyltransferases
There is only one chemically distinct structure:
Taxonomic group: bacteria / Firmicutes
(Phylum: Firmicutes)
The structure was elucidated in this paperNCBI PubMed ID: 27913418Publication DOI: 10.1128/AEM.02702-16Journal NLM ID: 7605801Publisher: American Society for Microbiology
Correspondence: simone.guglielmetti
unimi.it; a.p.laws
hud.ac.uk.
Institutions: Division of Food Microbiology and Bioprocesses, Department of Food, Environmental and Nutritional Sciences (DeFENS), Università degli Studi di Milano, Milan, Italy, Sofar S.p.A., Trezzano Rosa, Italy, Department of Biosciences, Università degli Studi di Milano, Milan, Italy, Department of Biological Sciences, University of Huddersfield, Queensgate, Huddersfield, United Kingdom
Lactobacillus paracasei DG is a bacterial strain with recognized probiotic properties and is used in commercial probiotic products. However, the mechanisms underlying its probiotic properties are mainly unknown. In this study, we tested the hypothesis that the ability of strain DG to interact with the host is at least partly associated with its ability to synthesize a surface-associated exopolysaccharide (EPS). Comparative genomics revealed the presence of putative EPS gene clusters in the DG genome; accordingly, EPS was isolated from the surface of the bacterium. A sample of the pure EPS from strain DG (DG-EPS), upon nuclear magnetic resonance (NMR) and chemical analyses, was shown to be a novel branched hetero-EPS with a repeat unit composed of l-rhamnose, d-galactose, and N-acetyl-d-galactosamine in a ratio of 4:1:1. Subsequently, we demonstrated that DG-EPS displays immunostimulating properties by enhancing the gene expression of the proinflammatory cytokines tumor necrosis factor alpha (TNF-α) and interleukin 6 (IL-6), and particularly that of the chemokines IL-8 and CCL20, in the human monocytic cell line THP-1. In contrast, the expression of the cyclooxygenase enzyme COX-2 was not affected. In conclusion, DG-EPS is a bacterial macromolecule with the ability to boost the immune system either as a secreted molecule released from the bacterium or as a capsular envelope on the bacterial cell wall. This study provides additional information about the mechanisms supporting the cross talk between L. paracasei DG and the host. IMPORTANCE: The consumption of food products and supplements called probiotics (i.e., containing live microbial cells) to potentially prevent or treat specific diseases is constantly gaining popularity. The lack of knowledge on the precise mechanisms supporting their potential health-promoting properties, however, greatly limits a more appropriate use of each single probiotic strain. In this context, we studied a well-known probiotic, Lactobacillus paracasei DG, in order to identify the constitutive molecules that can explain the documented health-promoting properties of this bacterium. We found a novel polysaccharide molecule, named DG-EPS, that is secreted by and covers the bacterium. We demonstrated that this molecule, which has a chemical structure never identified before, has immunostimulatory properties and therefore may contribute to the ability of the probiotic L. paracasei DG to interact with the immune system.
probiotic, MAMPs, Capsular EPS, Enterolactis, Lactobacillus casei DG, Lactobacillus paracasei, THP-1, immunostimulation
Structure type: polymer chemical repeating unit
Location inside paper: fig.3
Compound class: EPS
Contained glycoepitopes: IEDB_130648,IEDB_133754,IEDB_136044,IEDB_136105,IEDB_137472,IEDB_137473,IEDB_141794,IEDB_190606,IEDB_225177,IEDB_885823,SB_165,SB_166,SB_187,SB_195,SB_7,SB_88
Methods: 13C NMR, 1H NMR, NMR-2D, PCR, GC-MS, sugar analysis, DNA techniques, acid hydrolysis, GPC, extraction, permethylation, acetylation, cytokine production, bacterial adhesion to Caco-2 cells
Biological activity: EPS displays immunostimulatory properties by enhancing the gene expression of the proinflammatory cytokines TNF-α and IL-6 and, particularly, the chemokines IL-8 and CCL20.
Comments, role: published erroneous NMR chemical shift of #2,2,3,3,2_bDGalp С5 (70.0; matches aDGalp) was removed by CSDB staff
NCBI Taxonomy refs (TaxIDs): 1597Reference(s) to other database(s): GTC:G66763MN
Show glycosyltransferases
NMR conditions: in D2O at 343 K
[as TSV]
13C NMR data:
Linkage Residue C1 C2 C3 C4 C5 C6
2,2,3,3,2 bDGalp 106.0 72.5 76.5 70.1 ? 62.3
2,2,3,3 aLRhap 102.6 79.8 78.2 70.4 70.6 18.6
2,2,3 aLRhap 102.5 71.6 79.5 72.9 70.5 17.9
2,2,2 Ac 175.6 23.5
2,2 bDGalpN 103.5 57.1 83.1 72.6 77.3 62.4
2 aLRhap 102.4 80.3 71.3 73.7 73.8 18.0
aLRhap 102.2 79.6 72.0 73.6 70.6 18.1
1H NMR data:
Linkage Residue H1 H2 H3 H4 H5 H6
2,2,3,3,2 bDGalp 4.53 3.59 3.63 3.93 3.53 3.75
2,2,3,3 aLRhap 5.30 4.21 4.01 3.67 3.82 1.32
2,2,3 aLRhap 4.87 3.90 3.79 3.55 4.02 1.25
2,2,2 Ac - 2.05
2,2 bDGalpN 4.72 3.82 3.65 3.54 3.45 3.75-3.91
2 aLRhap 5.15 4.13 3.86 3.35 3.66 1.25
aLRhap 5.21 4.07 3.91 3.49 3.77 1.32
1H/13C HSQC data:
Linkage Residue C1/H1 C2/H2 C3/H3 C4/H4 C5/H5 C6/H6
2,2,3,3,2 bDGalp 106.0/4.53 72.5/3.59 76.5/3.63 70.1/3.93 ?/3.53 62.3/3.75
2,2,3,3 aLRhap 102.6/5.30 79.8/4.21 78.2/4.01 70.4/3.67 70.6/3.82 18.6/1.32
2,2,3 aLRhap 102.5/4.87 71.6/3.90 79.5/3.79 72.9/3.55 70.5/4.02 17.9/1.25
2,2,2 Ac 23.5/2.05
2,2 bDGalpN 103.5/4.72 57.1/3.82 83.1/3.65 72.6/3.54 77.3/3.45 62.4/3.75-3.91
2 aLRhap 102.4/5.15 80.3/4.13 71.3/3.86 73.7/3.35 73.8/3.66 18.0/1.25
aLRhap 102.2/5.21 79.6/4.07 72.0/3.91 73.6/3.49 70.6/3.77 18.1/1.32
1H NMR data:
| Linkage | Residue | H1 | H2 | H3 | H4 | H5 | H6 |
|---|
| 2,2,3,3,2 | bDGalp | 4.53 | 3.59 | 3.63 | 3.93 | 3.53 | 3.75 |
| 2,2,3,3 | aLRhap | 5.30 | 4.21 | 4.01 | 3.67 | 3.82 | 1.32 |
| 2,2,3 | aLRhap | 4.87 | 3.90 | 3.79 | 3.55 | 4.02 | 1.25 |
| 2,2,2 | Ac |
| 2.05 | |
| 2,2 | bDGalpN | 4.72 | 3.82 | 3.65 | 3.54 | 3.45 | 3.75
3.91 |
| 2 | aLRhap | 5.15 | 4.13 | 3.86 | 3.35 | 3.66 | 1.25 |
| | aLRhap | 5.21 | 4.07 | 3.91 | 3.49 | 3.77 | 1.32 |
|
13C NMR data:
| Linkage | Residue | C1 | C2 | C3 | C4 | C5 | C6 |
|---|
| 2,2,3,3,2 | bDGalp | 106.0 | 72.5 | 76.5 | 70.1 | ? | 62.3 |
| 2,2,3,3 | aLRhap | 102.6 | 79.8 | 78.2 | 70.4 | 70.6 | 18.6 |
| 2,2,3 | aLRhap | 102.5 | 71.6 | 79.5 | 72.9 | 70.5 | 17.9 |
| 2,2,2 | Ac | 175.6 | 23.5 | |
| 2,2 | bDGalpN | 103.5 | 57.1 | 83.1 | 72.6 | 77.3 | 62.4 |
| 2 | aLRhap | 102.4 | 80.3 | 71.3 | 73.7 | 73.8 | 18.0 |
| | aLRhap | 102.2 | 79.6 | 72.0 | 73.6 | 70.6 | 18.1 |
|
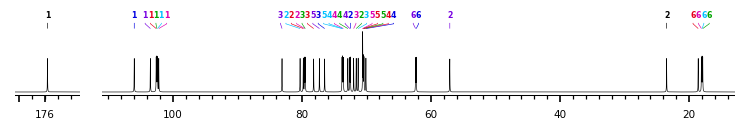
The spectrum also has 1 signal at unknown position (not plotted). |
There is only one chemically distinct structure:
 report error
report error Found 2 records.
Displayed records from 1 to 2
Found 2 records.
Displayed records from 1 to 2