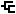Taxonomic group: bacteria / Proteobacteria
(Phylum: Proteobacteria)
The structure was elucidated in this paperNCBI PubMed ID: 34893256Publication DOI: 10.1016/j.carbpol.2021.118839Journal NLM ID: 8307156Publisher: Elsevier
Correspondence: R. Marchetti <roberta.marchetti
unina.it>; A. Silipo <silipo
unina.it>
Institutions: Department of Chemical Sciences and Task Force for Microbiome Studies, University of Naples Federico II, Via Cinthia 4, 80126 Naples, Italy, Department of Chemical, Materials and Production Engineering, University of Naples Federico II, Piazzale Tecchio 80, 80125, Naples, Italy, University of Cologne Cluster of Excellence on Plant Sciences (CEPLAS), Institute for Plant Sciences, D-50674 Cologne, Germany, ISIS Pulsed Neutron and Muon Source, Science and Technology Facilities Council, Rutherford Appleton Laboratory, Harwell Science and Innovation Campus, Didcot, Oxfordshire, OX11 OQX, UK, Instituto de Quimica Fisica Rocasolano, CSIC, Serrano 119, 28006 Madrid, Spain, CIBER de Enfermedades Respiratorias (CIBERES), Avda Monforte de Lemos 3-5, 28029 Madrid, Spain, Center for Proteomics and Metabolomics, Leiden University Medical Center, Leiden 2333 ZA, the Netherlands
Lipopolysaccharides, the major outer membrane components of Gram-negative bacteria, are crucial actors of the host-microbial dialogue. They can contribute to the establishment of either symbiosis or bacterial virulence, depending on the bacterial lifestyle. Plant microbiota shows great complexity, promotes plant health and growth and assures protection from pathogens. How plants perceive LPS from plant-associated bacteria and discriminate between beneficial and pathogenic microbes is an open and urgent question. Here, we report on the structure, conformation, membrane properties and immune recognition of LPS isolated from the Arabidopsis thaliana root microbiota member Herbaspirillum sp. Root189. The LPS consists of an O-methylated and variously acetylated D-rhamnose containing polysaccharide with a rather hydrophobic surface. Plant immunology studies in A. thaliana demonstrate that the native acetylated O-antigen shields the LPS from immune recognition whereas the O-deacylated one does not. These findings highlight the role of Herbaspirillum LPS within plant-microbial crosstalk, and how O-antigen modifications influence membrane properties and modulate LPS host recognition.
Lipopolysaccharide, NMR, Herbaspirillum, plant microbiota, structure-function relationship
Structure type: polymer chemical repeating unit
Location inside paper: Fig. 2, table S1
Compound class: O-antigen
Contained glycoepitopes: IEDB_1394181,IEDB_145006,IEDB_145010
Methods: 13C NMR, 1H NMR, NMR-2D, mild acid hydrolysis, MALDI-TOF MS, MD simulations, composition analysis, HPLC, molecular mechanics, microarray binding assays, DLS, SLS, elicitor preparation, chemiluminescence measurements, FTICR, specular neutron reflectometry measurements
Comments, role: NMR temperature was not specified; the published NMR assignment has many erroneous (impossible) chemical shifts of #3,4,3_aDRhap 100.69 73.05 79.80 71.00 69.40 16.70.
3D data: 3D data
NCBI Taxonomy refs (TaxIDs): 1890675
Show glycosyltransferases
NMR conditions: in D2O
[as TSV]
13C NMR data:
Linkage Residue C1 C2 C3 C4 C5 C6
3,4,3,3 Me 57.02
3,4,3 aDRhap
3,4,2 70%Ac 173.20 20.35
3,4 bDRhap 99.05 71.00 77.48 72.05 71.20 16.70
3,3 30%Ac 173.20 20.35
3 aDRhap 101.85 69.50 69.20 83.10 67.50 16.70
2 30%Ac 173.20 20.35
aDRhap 101.39 68.80 77.90 71.30 69.30 16.70
1H NMR data:
Linkage Residue H1 H2 H3 H4 H5 H6
3,4,3,3 Me 3.47
3,4,3 aDRhap 5.22 4.28 3.40 3.52 3.73 1.30
3,4,2 70%Ac - 2.22
3,4 bDRhap 5.00 5.50 3.89 3.57 3.55 1.37
3,3 30%Ac - 2.18
3 aDRhap 5.06 4.11 3.90 3.62 3.86 1.31
2 30%Ac - 2.18
aDRhap 4.91 4.09 3.82 3.56 3.73 1.28
1H/13C HSQC data:
Linkage Residue C1/H1 C2/H2 C3/H3 C4/H4 C5/H5 C6/H6
3,4,3,3 Me 57.02/3.47
3,4,3 aDRhap NMR TSV error 2: unequal length of 13C and 1H datasets
3,4,2 70%Ac 20.35/2.22
3,4 bDRhap 99.05/5.00 71.00/5.50 77.48/3.89 72.05/3.57 71.20/3.55 16.70/1.37
3,3 30%Ac 20.35/2.18
3 aDRhap 101.85/5.06 69.50/4.11 69.20/3.90 83.10/3.62 67.50/3.86 16.70/1.31
2 30%Ac 20.35/2.18
aDRhap 101.39/4.91 68.80/4.09 77.90/3.82 71.30/3.56 69.30/3.73 16.70/1.28
1H NMR data:
| Linkage | Residue | H1 | H2 | H3 | H4 | H5 | H6 |
|---|
| 3,4,3,3 | Me | 3.47 | |
| 3,4,3 | aDRhap | 5.22 | 4.28 | 3.40 | 3.52 | 3.73 | 1.30 |
| 3,4,2 | 70%Ac |
| 2.22 | |
| 3,4 | bDRhap | 5.00 | 5.50 | 3.89 | 3.57 | 3.55 | 1.37 |
| 3,3 | 30%Ac |
| 2.18 | |
| 3 | aDRhap | 5.06 | 4.11 | 3.90 | 3.62 | 3.86 | 1.31 |
| 2 | 30%Ac |
| 2.18 | |
| | aDRhap | 4.91 | 4.09 | 3.82 | 3.56 | 3.73 | 1.28 |
|
13C NMR data:
| Linkage | Residue | C1 | C2 | C3 | C4 | C5 | C6 |
|---|
| 3,4,3,3 | Me | 57.02 | |
| 3,4,3 | aDRhap | |
| 3,4,2 | 70%Ac | 173.20 | 20.35 | |
| 3,4 | bDRhap | 99.05 | 71.00 | 77.48 | 72.05 | 71.20 | 16.70 |
| 3,3 | 30%Ac | 173.20 | 20.35 | |
| 3 | aDRhap | 101.85 | 69.50 | 69.20 | 83.10 | 67.50 | 16.70 |
| 2 | 30%Ac | 173.20 | 20.35 | |
| | aDRhap | 101.39 | 68.80 | 77.90 | 71.30 | 69.30 | 16.70 |
|
There is only one chemically distinct structure:
 report error
report error Found 1 record.
Displayed record 1
Found 1 record.
Displayed record 1